
Structure determination of enzymes involved in osmolyte biosynthesis.
Structure determination of enzymes involved in osmolyte biosynthesis
In collaboration with the group of Prof. Dr. Erhard Bremer ( Univeristy of Marburg) try to elucidate the structures of enzymes/ protein involved in the biosynthesis of osmolytes. Here for example, ectoine and hydroxyectoine are well-recognized members of the compatible solutes and are widely employed by microorganisms as osmostress protectants. The EctABC enzymes catalyze the synthesis of ectoine from the precursor molecule L-aspartate-b-semialdehyde. A subgroup of the ectoine producers can convert ectoine into 5-hydroxyectoine through a regio-selective and stereospecific hydroxylation reaction. This compatible solute possesses stress-protective and function preserving properties different from those of ectoine. Hydroxylation of ectoine is carried out by the EctD protein, a member of the non-heme-containing iron(II) and 2-oxoglutarate-dependent dioxygenase superfamily. We were able to solve the structure of the EctD protein. This protein adopts a cupin fold and exhibits a iron ligand essential for catalysis. We determined the SaEctD crystal structure in its apo-form, complexed with the iron catalyst and in a form that contained iron, the co-substrate 2-oxoglutarate, and the reaction product of EctD, 5-hydroxyectoine.
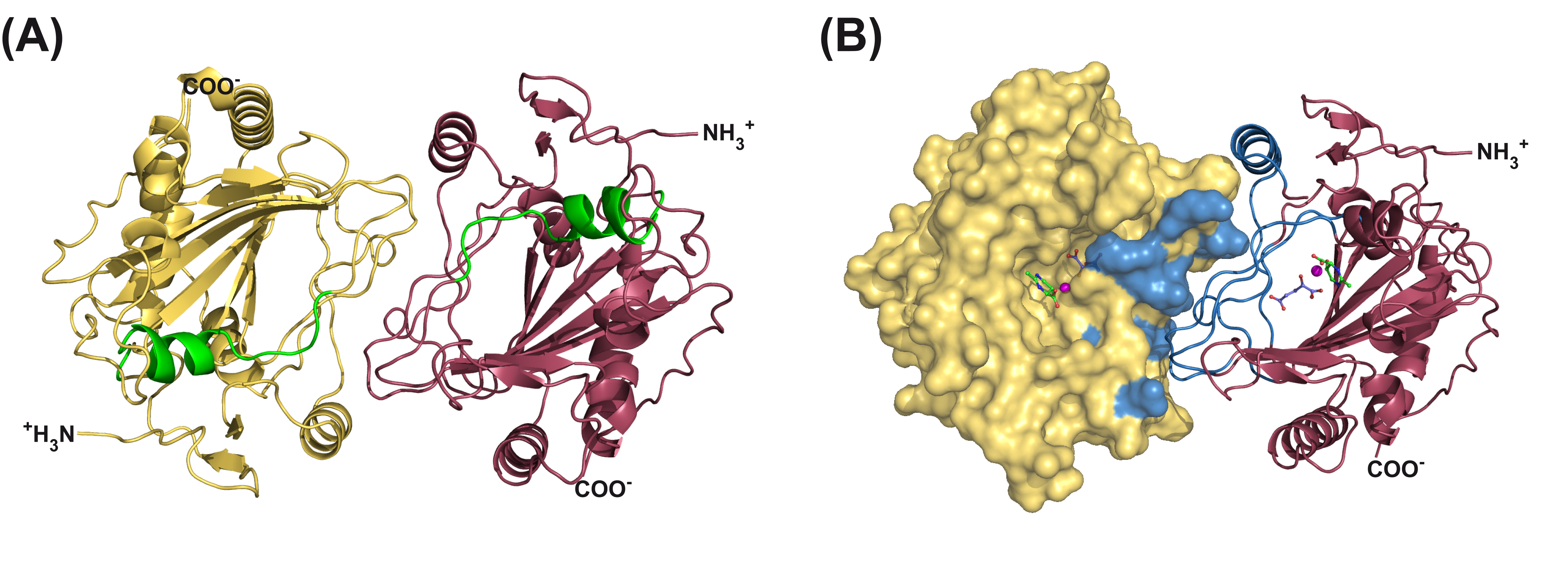
Figure 1. Structure of the SaEctD-Strep-tag-II protein in complex with iron, 2-oxoglutarate, and 5-hydroxyectoine.
The iron and 2-oxoglutarate ligands are bound within the EctD active site in a fashion similar to that found in other members of the dioxygenase superfamily. 5-hydroxyectoine on the other hand, is coordinated by EctD in manner different from that found in high-affinity solute receptor proteins operating in conjunction with microbial import systems for ectoines. Our crystallographic analysis provides a detailed view into the active site of the ectoine hydroxylase and exposes an intricate network of interactions between the enzyme and its ligands that collectively ensure the hydroxylation of the ectoine substrate in a position- and stereo-specific manner.
Recent publications
- Richter A.A., Kobus S., Czech L., Hoeppner A., Zarzycki J.Erb T.J., Lauterbach L., Dickschat J.S., Bremer E., Smits S.H. (2020) Making the versatile stress-protectant and chemical chaperone ectoine: architecture of the EctA diaminobutyrate acetyltransferase active site. JBC295 (9), 2822
- Hermann L., Mais C.N. Czech L., Smits S.H., Bange G., Bremer E. (2020) The Ups and Downs of Ectoine: Structural Enzymology of a Major Microbial Stress Protectant and Versatile Nutrient accepted in Biol Chem
- Czech L., Hoeppner A., Smits S.H., Bremer E. (2020) Ectoine synthase: an iron-dependent member of the cupin superfamily Encyclopedia of Inorganic and Bioinorganic Chemistry DOI: 10.1002/9781119951438.eibc2742
-
Czech L., Höppner A., Kobus S., Seubert A., Riclea R., Dickschat J.S., Heider J., Smits S.H., and Bremer E. (2019) Illuminating the catalytic core of ectoine synthase through structural and biochemical analysis Scientific Reports 9:364 | DOI:10.1038/s41598-018-36247-
-
Czech L., Wilcken S., Czech O., Linne U., Brauner J., Smits S.H., Galinski E.A., Bremer E. (2019) Exploiting substrate promiscuity of ectoine hydroxylase for regio-and stereoselective modification of homoectoine Frontiers in Microbiology 10:2745. doi: 10.3389/fmicb.2019.02745
-
Richter A.A., Mais C-N., Czech L., Geyer K., Hoeppner, A. Smits S.H., Erb T.J.,Bange G., Bremer E. (2019) Biosynthesis of the stress-protectant and chemical chaperon ectoine: biochemistry of the transaminase EctB Frontiers in Microbiology 10:2811. doi: 10.3389/fmicb.2019.02811
- Czech L., Hermann L., Stöveken N., Richter AA., Höppner A., Smits, SH, Heider J., and Bremer E. (2018) Role of the extremolytes ectoine and hydroxyectoine as both stress protectants and nutrients: genetics, phylogenomics, biochemistry, and structural analysis Gene 210.432
- N. Widderich, E. Bremer, S.H.J. Smits The ectoine hydroxylase: a non-heme-containing iron(II) and 2-oxoglutarate-dependent dioxygenase (2016) Encyclopedia of Inorganic and Bioinorganic Chemistry chapter 1
- Widderich, N., Kobus, S., Hoeppner, A., Riclea, R., Seubert, A., Dickschat, J.S., Heider, J., Smits, S.H.J., Bremer, E., Biochemistry and crystal structure of ectoine synthase: an iron-containing member of the cupin superfamily PLoS ONE 11(3): e0151285. doi:10.1371/journal.pone.0151285
- Broy S, Chen C, Hoffmann T, Brock NL, Nau-Wagner G, Jebbar M, Smits SH, Dickschat JS, Bremer E. (2015 ) Abiotic stress protection by ecologically abundant dimethylsulfoniopropionate and its natural and synthetic derivatives: insights from Bacillus subtilis. Environ Microbiol. 17, 2362-78
- Widderich, N., Pittelkow, M., Hoppner, A., Mulnaes, D., Buckel, W., Gohlke, H., Smits, S. H., and Bremer, E. (2014) Molecular dynamics simulations and structure-guided mutagenesis provide insight into the architecture of the catalytic core of the ectoine hydroxylase. Journal of molecular biology 426, 586-600
- Widderich, N., Hoppner, A., Pittelkow, M., Heider, J., Smits, S. H., and Bremer, E. (2014) Biochemical properties of ectoine hydroxylases from extremophiles and their wider taxonomic distribution among microorganisms. PloS one 9, e93801
- Hoeppner A, Widderich N, Lenders M, Bremer E, Smits SH (2014) Crystal structure of the ectoine hydroxylase, a snapshot of the active site. J Biol Chem 289: 29570-29583
- Bashir, A., Hoffmann, T., Smits, S. H., and Bremer, E. (2014) Dimethylglycine provides salt and temperature stress protection to Bacillus subtilis. Applied and environmental microbiology 80, 2773-2785
- Bashir A, Hoffmann T, Kempf B, Xie X, Smits SH, et al. (2014) Plant-derived compatible solutes proline betaine and betonicine confer enhanced osmotic and temperature stress tolerance to Bacillus subtilis. Microbiology 160: 2283-2294